BIRD NAVIGATION EXPERIMENTS
Extract from "Bird Migration" by Donald
R. Giffin
While the evidence summarized in the previous
chapter did suggest that herring gulls and gannets managed to home without
any highly developed ability to select the home direction, there was no
compelling reason to apply this explanation to all birds. Suspecting that
other kinds of birds might well display superior powers of navigation,
Harold B. Hitchcock and I turned to homing pigeons in an attempt to learn
something about their navigation by tracing their flight paths from small
airplanes. Hitchcock's pigeons performed better than mine, and therefore
Fig. 16 shows some of the longer flight paths which he succeeded in following.
It is apparent that these pigeons deviated considerably from the direct
path between release point and home, but they did better than one would
expect from the molecular theory of bird orientation.
In the early 1950s two important advances
were achieved in the explanation of bird navigation, one by Geoffrey V.
T. Matthews at Cambridge University, in England, the other by Gustav Kramer
at Wilhelmshaven, Germany. Matthews worked mainly by observing and analyzing
the initial headings of birds in homing experiments. Kramer's experiments
(to be discussed later) included not only observation of the initial headings
of released homing pigeons but also study of the migration restlessness
(described in Chapter 3) of a few selected, handraised, and caged wild
birds.
For many years pigeon racers had maintained
that on release homing pigeons often would fly straight off toward home.
But in most casess racing pigeons are released in large flocks containing
many birds experienced withh the particular release point and the homeward
route. A reasonable explanation, in the absence of other information, was
that these experienced birds remembered the correct route and were followed
by the others. In many homing experiments the immediate surroundings at
the release point sometimes seemed to have an effect on the bird's choice
of direction for its first flight. For instance, sea birds released inland
tended to head for any body of water, while pigeons were likely to display
some interest in towns or farms.
Matthews' Observations of Initial Headings
Matthews made a special effort to select
release points at the centers of large open spaces of level terrain with
a clear view in all directions. Every effort was made to avoid local features
of topography that might influence the direction in which the birds would
fly when first set free. To guard against bias it was necessary, of course,
to vary the direction in which the birds' heads were pointed at the actual
moment of release. This pointing was done in an irregular fashion, perhaps
with the first bird headed north, the next southwest, the third east, and
so forth.
In his first experiments Matthews trained
pigeons by releasing them at increasingly distant points that all lay approximately
on a single line. This training direction extended north northwest from
Cambridge. As in all properly conducted homing experiments, the birds were
transported in closed boxes, usually by automobile or light truck, and
they had no opportunity to see anything of the countryside. When thus released
in unknown te.ritory in an unknown direction from home, many of the pigeons
headed initially in their customary southward homing direction. These birds,
it was apparent, had established a preferred direction of flight which
they chose regardless of the release point. When taken to a new release
point on the training line, several miles farther north than their previous
starting point, the pigeons again displayed a clear ability to fly south-toward
Cambridge.
Naturally, Matthews wanted to find out
on what basis this choice was made. By releasing pigeons under a variety
of weather conditions, he made the important discovery that the initial
headings were far less consistent when the sky was heavily enough overcast
to hide the sun. While there was the usual variahility from day to day,
and from one pigeon to another, the choice of an appropriate direction
seemed to be greatly impeded, if not rendered altogether impossible, when
the pigeon could not see the sun. It had long ago been suggested that in
pigeon races, which always start early in the morning, the birds inight
take advantage of the sun's position to deterinine the appropriate homeward
direction in which to fly. But it seemed incredible that a bird might use
the sun as a "compass" to determine directions at any time of day. Matthews
was the first to consider this possibility seriously enough to carry out
esperijnents that tested it directly.
In one series of releases along in the
training diicction north northwest from the pigeon's home aloft near Cambridge
the releases were made always iii the middle of the day when the sun was
nearly at its highest position in the sky. It was thus approxiiiately south
of the birds when they made their ini( iA choices of flight direction.
The simplest esplanai ion for these directional choices would be that the
I)ird had learned to fly toward the sun. But these pi;cons did not merely
start in the correct direction; niany of them also reached home at relatively
high speeds. A simple tendency to head toward the sun would have caused
them to turn more and more to the west as the afternoon wore on, and by
evening they would have been flying almost directly away from home.
Matthews investigated the matter further
by experimenting with pigeons which had been released several times at
increasing distances up to seventyeight miles along the training line,
always at approximately midday. Dividing the flock into three groups, he
transported them forty-nine miles farther (that is, 127 miles from home)
in the same direction. One group was now released early in the morning,
another late in the afternoon, and a control group in the middle of the
day as usual. On the average the experimentals were released six hours
earlier or later than the controls. The result was that all three groups
chose the homeward direction with equal accuracy. It made no difference
that the sun was roughly ninety degrees east or west of the position where
they might have come to expect it. The results of this important experiment
are illustrated in Fig. 17, which shows not only the similarity of the
headings when birds were released early in the morning, at midday, and
late in the afternoon, but also the general accuracy of these homeward
headings.
After Matthews had completed the experiments
just described, he did not rest content with the demonstration that his
pigeons could use sun-position for headings. He continued and extended
the same basic type of experiment and soon found that some groups of pigeons
performed in a still more interesting manner. After considerable experience
at distances up to fifty miles or more, along a single training line, they
were released in unknown territory in some quite different direction. In
the most important experiments the training direction was again north northwest,
but the crucial tests were made at release points to the south or west.
Again there was considerable variability, and a few of the birds flew roughly
south as they had done on previous occasions when released along the training
line. Others scattered in many other directions. But a considerable majority
flew off more or less toward home. This was true whether home was north
or east. After a number of experiments had been completed, it became clear
that many of the pigeons could select approximately the homeward direction
whether or not it coincided with that of the previous training flights.
Examples of several experiments of this type are shown in Fig. 18. Despite
the considerable variability, the choices made by the great majority of
birds lay far too close to the home direction to be explained by random
scattering, or by any of the explanations that appeared applicable to the
gannets and other homing experiments prior to those of Matthews.
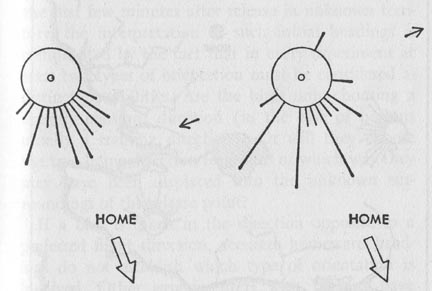
Fig. 17, Initial headings of pigeons
trained by several releases at midday. Each release was farther north northwest
from the home loft, the release previous to this one having been at 78
miles. At left, the initial headings of birds released at 127 miles (in
totally strange territory) around noon. At right, the headings of others
from the same group released at the same place in early morning and late
afternoon. The length of each line is proportional to the number of pigeons
starting in the direction indicated; the shortest lines signify one bird.
Small arrows show the directions these birds would be expected to take
if they had flown at the same angle to the sun as in the training flights.
In spite of the definite demonstration
that pigeons could establish homeward orientation within the first few
minutes after release in unknown territory, the interpretation of such
initial headings is complicated by the fact that in every experiment at
least two types of orientation must be considered as distinct possibilities.
Are the birds only choosing a roughly constant direction (in the case of
pigeons usually a training direction)? Or will they choose the true home
direction regardless of which way they may have been displaced into the
unknown surroundings of the release point?
If a bird is taken in the direction opposite
to a preferred flight direction, accurate homeward headings do not establish
which type of orientation is involved. Other experimenters with pigeons
have found that their birds exhibit a constant directional tendency in
their initial headings independent of the direction they have been carried
from home. For example, pigeons trained to home to Wilhelmshaven near the
northern coast of Germany, where most of Kramer's experiments were conducted,
show a northward tendency in their initial headings. Only after releases
in two or more different directions can one determine with reasonable confidence
whether a uni- directional tendency is at work or true homeward orientation.
The Initial Headings of Manx Shearwaters
In addition to his work with domestic homing
pigeons, Matthews also experimented with Manx shearwaters, birds which
had already been shown to have a most impressive homing ability. It was
Manx shearwaters which returned from Venice and Boston to their nesting
island off the coast of Wales at average speeds of 265 and 244 miles per
day. Matthews transported shearwaters from the same island to various points
inland in England where the terrain was relatively uniform and an observer
on a good vantage point (such as the library tower at Cambridge University)
could follow the birds with binoculars for considerable distances. Again
there was variability, some shearwaters choosing directions far removed
from the correct homeward path. But, on the average, shearwaters performed
as well or better than the best homing pigeons, and a tabulation of results
from several inland release points in England showed beyond doubt that
most of the shearwaters were choosing approximately the correct homeward
direction and were not scattering in anything approaching a random pattern.
These experiments obviously carried the
whole problem of homing in birds into a new phase. Other investigators
tried essentially the same methods on other species, but no other bird
has yet been found which shows as consistently accurate initial headings
as the homing pigeons, used by Matthews and Kramer, or the Manx shearwater.
Other not-so-select strains of homing pigeons give results that are far
less convincing. Nevertheless, the fact that some birds can consistently
choose approximately the correct homeward direction, and demonstrate the
correctness of this choice in their initial headings, means that we are
dealing with an ability to orient which makes itself manifest within a
very short time after release. No one can yet say with certainty whether
it is only a few species of wild birds, and only the best strains of homing
pigeon, that possess this ability, or whether it is widespread among birds
but has only been demonstrated in a few favorable cases.
Kramer's Experiments with Orientation
Cages
At about the time when Matthews was conducting
the experiments described, Gustav Kramer and his associates were experimenting
with orientation exhibited under more strictly controlled conditions in
experimental orientation cages. In his first investigations Kramer used
starlings which had been raised by hand from an early age, and were very
tame. They were placed in circular cages provided with perches both at
the center and also around the edges. As had been discovered many years
earlier, such migratory birds exhibit a pronounced restlessness at the
season when they would normally undertake long migratory flights. As explained
at the end of Chapter 3, this migratory restlessness occurs primarily at
night, when many small birds migrate, but Kramer's clearest experiments
were carried out in daylight, employing a few well-tamed starlings, which
are diurnal migrants.
The starlings were placed in a circular
cage which was carefully built so as to be completely symmetrical; its
inner surface had an identical appearance when the bird faced in any direction.
The observer lay on his back on the floor and watched the birds from
below through the transparent plastic floor of the orientation cage. One
tame and co-operative starling tended in the spring to fly back and forth
from the central perch to the edge of the cage primarily in a northwesterly
direction. This direction corresponded roughly to the normal northeasterly
spring migration of starlings in the area where it had been taken from
its nest in its first few days of life.
On what basis did this starling make its
directional choice? In addition to being symmetrical, the orientation cage
was arranged so that it could be rotated, and when it was turned from time
to time during the period of migratory restlessness the starling continued
to head in the same general direction, regardless of which wall of its
cage had to be approached in order to head northwest from the central perch.
The next step was to surround the orientation cage with a uniform, opaque
fence or shield, which also could be rotated about the center of the cage.
This screen cut off any view of the surrounding landmarks, but allowed
the starling to see most of the sky. Again it made no difference how the
screen was rotated. Clearly the bird was not responding to local landmarks.
Kramer also established that the bird and orientation cage could be moved
about from point to point in the vicinity of Wilhelmshaven, without occasioning
any essential change in the directional tendency.
What aspect of the sky provided the directional
information underlying these choices of the correct direction for fall
migration? One possible directional cue was the sun. For the decisive experiment
Kramer enclosed the orientation cage in a further opaque structure which
excluded all view of the outside world except what the bird could see through
six large windows. This starling continued to show the same northwesterly
heading when it could see the sun and blue sky through one or more of these
windows. The next step was to equip the windows with shutters whose inner
surfaces had large plane mirrors, as shown in Fig. Iq. These shutters were
so arranged that the light reflected into the window by the mirror came
from ninety degrees to the right or left of the window itself. In other
words, looking out the window, the bird saw reflected in the mirror an
area of sky ninety degrees to one side of the sky directly beyond the window.
The starling continued to show strong directional preferences, but now
the bird maintained the same angle relative to the light reflected from
the mirrors as it bad to the normal sunlight. Thus, when the mirrors showed
the sky ninety degrees to the right of what normally would have been visible
through each window, the starling headed approximately ninety degrees to
the right of its previous heading. The opposite result occurred when the
windows were so arranged that the sunlight reached the bird from ninety
degrees to the left of its normal angle of incidence.
When the sky was completely overcast the
headings were virtually random, but when the sun became visible a few hours
later the bird again chose approximately the northwest direction for its
headings. Kramer thus showed that one particularly cooperative starling
was able to choose a consistent direction in his circular orientation cage,
and, furthermore, that this direction was determined with reference to
the sun. The correspondence between this experiment and Matthews' studies
of initial flight direction in pigeons was obvious and compelling.
ANIMAL NAVIGATION
(Extract from Animal Navigation by R. M. Lockley Science Series,
1967)
It is now known that many if not most of
the non-migratory birds lack - presumably because they have never had occasion
to exercise it - the innate ability to orientate over long distances possessed
by pelagic and oceanic migrants such as the albatrosses, shearwaters, terns,
etc. At most the purely sedentary species can get back home only if they
are released within or close to their local territories, with some exceptional
individuals doing rather better than this. The case of the racing, carrier
or homing pigeon is interesting, because, although it has been extensively
used in scientific homing tests, it is also by origin a non-migratory bird.
It derives ftom. the wild rock-dove, normally resident within a very few
miles of its rock caves and cliffs throughout the year - a strictly vegetarian
feeder.
Tales of prodigious horming feats of this
domesticated dove must be discounted; they may be boasts of imaginative
racing pigeon fanciers. The many careful experiments prove that their sense
of direction is limited, and that they are lost if transported hundreds
of miles out of the area in which they have been trained to race. Like
dogs which have been trained to home in a specific direction they will
return home rapidly over lengthening distances only if released in strange
country along the compass course with which they are familiar. Even so
it is usually necessary for pigeons to see the sun (they do not fly at
night) so as to orient on the correct bearing for home. Racing pigeon fanciers
fear misty weather when they send their trained birds away on competitive
races; many are lost then, wandering to strange lofts, or settling down
to live with wild rock-doves on cliffs, or gone-wild pigeons on town buildings.
They seem to lose heart if they cannot get back to their home loft in the
same day; and one pigeon fancier tells me that even if he does eventually
recover a lost bird it is seldom much use as a racer afterwards and he
will not keep it as a breeder in case it perpetuates its failure in its
offspring.
In a series of experiments which I conducted
with the Editor of the Racing Pigeon, five untrained young pigeons
were released in Wales 250 miles from their home loft in London, and five
more from the same loft at Cardiff, i5o miles from London. Although these
youngsters had been flying strongly above their London loft, and knew it
visually from the air, they had never been sent away on races, but were
robust and physically in perfect flying condition. Yet none was ever recovered
back home again; only two were reported, some weeks later, at lofts completely
off course in the Western Midlands, at least 70 miles from London. This
proved that these young domesticated rock doves, like other non-migratory
birds, have no innate ability to navigate when displaced so far out of
their normal range. They are convenient to use because of their sedentary
habit of returning to a communal roost, but not ideal navigators in such
experiments.
It is, however, possible to train them,
and it was with well trained pigeons that G. V. T. Matthews had some interesting
results. By releasing groups of experienced homers farther along their
training line at different hours of the day, he found that the position
of the sun in the sky at the moment of release made no difference: the
majority flew off in the correct direction for home, and only a minority
headed in the opposite direction.
The experiments were repeated on a larger
scale, with five widely separate release points; this time many of the
birds were released in unknown country in a direction opposite to that
of the previous training one. The results are shown in Figure 10, and confirm
that some, but not all, pigeons are able, after training, to select a homeward
direction if released almost anywhere within i5o miles of home. But in
misty weather they were quite disoriented.
The homing experiments with oceanic birds
quoted above indicate that those species, under test, can correct much
longer deviations from their normal range artificially arranged by man,
and find their way home over many hundreds of miles of unknown sea and
land. On reflection it is clear that such an ability is essential for survival
when strong winds blow individuals far off course into regions unfamiliar
to them, but how they re-orient so accurately remains a puzzle.
Several years ago G. V. T. Matthews
suggested that the day-flying homing pigeon at the moment of release is
able to detect the arc through which the sun is travelling, and predict
the arc to its highest point (due south in the northern hemisphere) and
so obtain its latitude. At the same moment the bird's keen sense of time
tells it where the sun ought to be at home (or home time at noon) which
will give it its relative longitude. But how precise a bird can be in calculating
so fine a distinction we do not know. When man makes such a calculation
by the heavenly bodies he has to allow for several, but principally three,
variations: for dip, which is the angle of epression of the visible
below the sensible horizon; for parallax, the angle subtended at the heavenly
body's centre by the observer and the earth's centre (when a body is at
the observer's zenith the angle is 0 degrees); and especially refraction
- a ray of light from a celestial body is bent as it passes through layers
of increasing density of the earth's atmosphere.
In the northern hemisphere mallard, in
homing tests in both England and the United States, seem to be drawn towards
the North Star in what Matthews describes as 'nonsense orientation', before
they eventually swing round and resume their migration in the normal direction.
In eastern North America Grfffin found that common terns headed initially
in the opposite direction - south cast; although some of the terns were
nesting in that direction, others were breeding north-east of the release
point. Even more puzzling is the report that some common and black-headed
gulls were able to orient themselves towards the place of capture from
a totally enclosed room with no view of the sky.
Matthews's 'North Star mallards' were the
subject of several other experiments, continuing as I write this. He 'reset'
their biological clocks by keeping them away from daylight in an enclosure,
under artificial schedules of light, for several days until some were accustomed
to a day six hours ahead, and some to a day six hours behind, the true
day. A third group was accustomed to a day twelve hours out of phase. When
he released in daylight all three groups, together with a control group
which had been living under natural daylight, the controls headed north-west
as expected. Those with clocks reset six hours ahead flew south-west, those
with clocks six hours slow flew north-east, and those with clocks twelve
hours out of phase, flew in the opposite direction - south-east. Thus the
experiment with sandhoppers was duplicated with ducks, with the same
result. But when Matthews released the same three groups of ducks with
clocks reset as before, but under the stars on a clear night, all the mallard
oriented northwest, as did the controls!
Man has found that the stars (but not the
planets), fixed in their courses at an immense distance from Earth, are
more reliable as navigation guides than the great sun itself. So, too,
it seems, sidereal pattern and time have a greater influence on animal
navigation than the sun, at least where a species, like the mallard, migrates
by both day and night.
There is an acceptable theory that in general
migrating birds follow the shortest possible line between summer and winter
quarters, a line which can only be drawn accurately on a globe atlas. Any
'straight' line extended far enough over and parallel to the surface of
the earth will automatically complete a circle of the same length as the
equator. Such a line is known as a Great Circle. Plotted on a flat projection
of the world, the flight of a bird such as a warbler, migrating from summer
quarters in eastern Siberia straight to winter quarters in Kenya, would
appear to pass southwards through the Gobi Desert of China, India, and
over the western Indian Ocean. In fact its shortest route along the great
circle actually begins with a westward flight into Europe, and only gradually
southward via the Aral Sea and the Red Sea to Kenya - an all land-route
and less perilous for a small insectivorous warbler.